

Três características e funções no organismo
- 3326
- 339
- Shawn Leffler
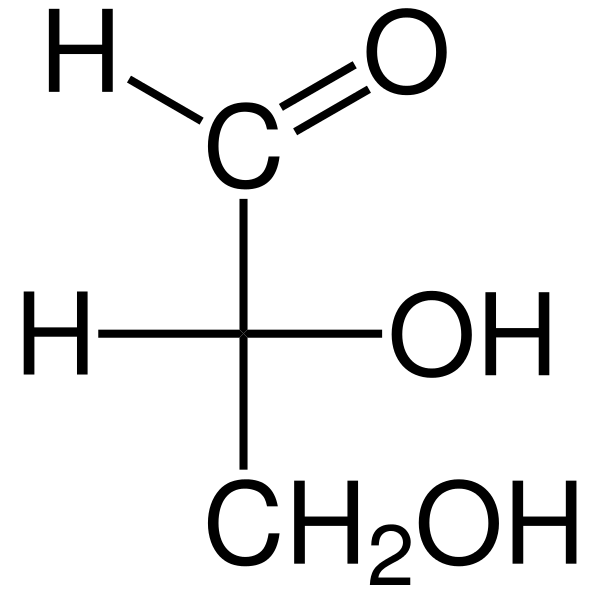
As Triosas São monossacarídeos de três carbonos cuja fórmula química empírica é C3H6QUALQUER6. Existem dois triosas: gliceraldehyd. Os triosas são importantes no metabolismo porque conectam três vias metabólicas: glicólise, gliconeogênese e o caminho de fosfato de pentose.
Durante a fotossíntese, o ciclo Calvin é uma fonte de triosas que servem para a biossíntese de frutose-6-fosfato. Este açúcar, de maneira fosforilada, é convertida por etapas enzimaticamente catalisadas em polissacarídeos de reserva ou estrutura estrutural.
Fonte: Wesalius [domínio público] Os triosas participam da biossíntese de lipídios que fazem parte de membranas celulares e adipócitos.
[TOC]
Caracteristicas
O gliceraldeído aldosa possui um átomo de carbono quiral e, portanto, possui dois enantiomeros, o L-Gliceraldeído e o D-Glérico. Ambos os enantiômeros d e l têm diferentes características químicas e físicas.
O D-gliceraldeído quebrou o plano da luz polarizada à direita (+) e tem uma rotação [α] D, a 25 ° C, de +8.7 °, enquanto o L-gliceraldeído quebrou o plano da luz polarizada à esquerda (-) e tem uma rotação [α] D, a 25 ° C, de -8.7 °.
O carbono quiral de gliceraldeído é carbono 2 (c-2), que é um álcool secundário. A projeção de Fischer representa o grupo hidroxila (-OH) do D-gliceraldeído à direita e o grupo OH- do L-Glyce esquerdo.
A di -hidroxiacetona carece de carbonos quirais e não possui formas enantioméricas. A adição de um grupo hidroximetileno (-CHOH) a gliceraldeído ou di-hidroxiacetona permite a criação de um novo centro quiral. Consequentemente, o açúcar é uma tetrosa porque tem quatro carbonos.
A adição de um grupo -Chah a Terosa cria um novo centro quiral. O açúcar formado é um pentose. Grupos -Chah pode continuar a ser adicionado a um máximo de dez carbonos.
Pode atendê -lo: filogeniaFunções no organismo
Os triosas como intermediários em glicólise, gluconeogênese e o caminho de fosfato de Pentosa
A glicólise consiste na ruptura da molécula de glicose em duas moléculas de piruvato para produzir energia. Esta rota implica duas fases: 1) fase preparatória ou consumo de energia; 2) fase de geração de energia. O primeiro é o que produz os triosas.
Na primeira fase, o teor de energia livre de glicose é aumentado, pela formação de fosfoésters. Nesta fase, o trifosfato de adenosina (ATP) é o doador de fosfato. Essa fase culmina com a conversão de frutose Phosfoéster 1.6-bifosfato (F1.6bp) em dois trisasas fosfato, gliceraldeído 3-fosfato (GA3p) e di-hidroxiacetona-fosfato (DHAP).
A gliconeogênese é a biossíntese de glicose do piruvato e outros intermediários. Use todas as enzimas de glicólise que catalisam as reações cuja variação de energia padrão bioquímica está em equilíbrio (ΔGº '~ 0). Por causa disso, a glicólise e a gliconeogênese têm intermediários comuns, incluindo GA3P e DHAP.
O caminho da pentose fosfato consiste em dois estágios: uma fase oxidativa de glicose-6-fosfato e outra formação de NADPH e ribosa-5-fosfato. Na segunda fase, a ribose de 5-fosfato é convertida em intermediários de glicólise, F1.6bp e Ga3p.
Os triosas e o ciclo Calvin
A fotossíntese é dividida em dois estágios. No primeiro, ocorrem reações dependentes da luz que produzem NADPH e ATP. Essas substâncias são usadas no segundo, na qual há fixação de dióxido de carbono e formação de hexose a partir de triosas através de um caminho conhecido como Calvin Cycle.
No ciclo Calvin, a enzima de 1,5-bifary-oxigenase (Rubisco) catalisa a união covalente do CO2 Para a pentose ribulosa 1.5-bifosfato e quebra o intermediário instável de seis átomos de carbono em duas moléculas de três átomos de carbono: o 3-fosfoglicerado.
Pode atendê -lo: oligossacarídeos: características, composição, funções, tiposAtravés de reações enzimáticas que incluem fosforilação e redução do 3-fosfoglicerado, usando ATP e NADP, ocorre Ga3p. Este metabolito é convertido em 1,6-bifosfato frutose (F1.6bp) por uma via metabólica semelhante à gliconeogênese.
Por ação de uma fosfatase, a F1.6bp é convertida em frutose-6-fosfato. Em seguida, uma fosfhexose de isoma produz glicose 6-fosfato (GLC6P). Finalmente, uma epicherase converte Glc6p em glicose 1-fosfato, que serve para a biossíntese de amido.
As triosas e lipídios de membranas biológicas e de adipócitos
GA3P e DHAP podem formar fosfato glicerol, que é um metabolito necessário para a biossíntese de triacilgliceroles e glicerolipídios. Isso ocorre porque ambos os fosfatos de triosas podem ser interconvertidos por uma reação catalisada pelo fosfato de triosa isomerase, que mantém os dois triosas em equilíbrio.
A enzima de glicerol-fosfato desidrogenase catalisa uma reação de redução de oxid, na qual o NADH doa alguns elétrons ao DHAP para formar glicerol de 3-fosfato e NAD+. L-glicerol 3-fosfato faz parte do esqueleto de fosfolipídios que são parte estrutural das membranas biológicas.
O glicerol é pró-renda, não possui carbonos assimétricos, mas quando um de seus dois álcoois primários forma um fosfoester, ele pode ser chamado corretamente.
Os glicofosfolipídios também são chamados de fosfoglicerídeos, sendo nomeados como ácido fosfatídico derivado. Os fosfoglicerídeos podem formar fosfoacilgliceroles formando ligações éster com dois ácidos graxos. Nesse caso, o produto resultante é de 1,2-fosfodiacilglicerol, que é um componente importante das membranas.
Uma glicofosfase catalisa a hidrólise do grupo fosfato de glicerol de 3-fosfato, produzindo glicerol mais fosfato. O glicerol pode servir como um metabólito inicial para a biossíntese de triacilglicerídeos, que são comuns em adipócitos.
Pode atendê -lo: teste de coagulase: fundação, procedimento e usosAs triosas e membranas das arqueobactérias
Semelhante a eubactérias e eucariotos, o glicerol de 3-fosfato é formado a partir de fosfato de triosas (GA3P e DHAP). No entanto, existem diferenças: a primeira é que o glicerol de 3-fosfato nas membranas de arqueobactérias é a configuração de L, enquanto nas membranas eucarióticas Eubactérias e eucarióticas é a configuração D.
Uma segunda diferença é que as membranas dos arqueobactérias formam éster vincula-se a duas longas cadeias de hidrocarbonetos de grupos isoprenóides, enquanto em eubactérias e eucariotos, o glicerol forma ligações éster (1.2-diacilglicerol) com duas cadeias de hidrocarbonetos de ácidos graxos.
Uma terceira diferença é que, nas membranas dos arqueobactérias, as substituições no fosfato e no glicerol 3-fosfato são diferentes das de eubactérias e eucariotos. Por exemplo, o grupo fosfato está ligado ao dissacarídeo α-Glucopiraano- (1®2)-β-Galatofuranosa.
Referências
- Cui, s. C. 2005. Carboidratos de alimentos: química, propriedades físicas e aplicações. CRC Press, Boca Raton.
- Cock, p., Mäkinen, K, Honkala e., Saag, m., Kennepohl, e., Eapen, a. 2016. O eritritol é mais eficaz que o xilitol e o sorbitol no gerenciamento de terminais de saúde bucal. Jornal Internacional de Odontologia.
- Nelson, d. eu., Cox, m. M. 2017. Lehninger Principles of Biochemistry. C. H. Freeman, Nova York.
- Sinnott, m. eu. 2007. Química de carboidratos e estrutura e mecanismo de bioquímica. Sociedade Real de Química, Cambridge.
- Stick, r. V., Williams, s. J. 2009. Carboidratos: as moléculas essenciais da vida. Elsevier, Amsterdã.
- Voet, d., Voet, J. G., Pratt, c. C. 2008. Fundamentos da Bioquímica - Vida no nível molecular. Wiley, Hoboken.
- « Características da aldose, tipos, número de carbonos
- Mecanismo de ação e efeitos colaterais da fluoxetina (Prozac) »