

Características de poros nucleares, funções, componentes
- 3925
- 479
- Lonnie MacGyver
Ele poro nuclear (Do grego, poros = etapa ou trânsito) é a "porta" do núcleo que permite a maior parte do transporte entre nucleoplasma e citoplasma. O poro nuclear se junta às membranas internas e externas do núcleo para formar canais, que servem para o transporte de proteínas e RNA.
A palavra Poro não reflete a complexidade da estrutura em questão. Por causa disso, é preferível se referir ao complexo de poros nucleares (CPN), em vez de poro nuclear. A CPN pode experimentar mudanças em sua estrutura durante o transporte ou estados do ciclo celular.
Recentemente, descobriu -se que nucleoporinas, proteínas que compõem o CPN, têm um papel importante na regulação da expressão gênica. Assim, quando ocorrem mutações que afetam a função das nucleoporinas, as patologias são produzidas em humanos, como autoimunes, cardiomiopatias, infecções virais e câncer.
[TOC]
Caracteristicas
Através da tomografia eletrônica, foi determinado que o CPN tem uma espessura de 50 nm, um diâmetro externo entre 80 e 120 nm e um diâmetro interno de 40 nm. Grandes moléculas como a grande subunidade de ribossomos (PM 1590 kDa) podem ser exportadas para fora do núcleo através do CPN. Estima -se que existem entre 2000 e 4000 cpn por núcleo.
O peso molecular de um CPN individual é de aproximadamente 120 e 125 MDA (1 MDA = 106 Da) em vertebrados. Por outro lado, o CPN é menor em leveduras, nas quais possui aproximadamente 60 mdA. Apesar do enorme tamanho da CPN, as nucleoporinas são altamente preservadas em todos os eucariotos.
A translocalização através do CPN é um processo rápido, cuja velocidade é 1000 translocações/segundo. No entanto, o CPN não determina a direção do próprio fluxo de transporte.
Isso depende do gradiente Rangtp, que é maior no núcleo do que no citosol. Este gradiente é mantido por um fator de trocador de guanina executado.
Durante o ciclo celular, os CPNs experimentam ciclos de montagem e desunião. A montagem acontece na interface e imediatamente após a mitose.
Funções
Ácido ribonucleico (pequeno RNA nuclear, RNA mensageiro, RNA de transferência), proteínas e ribonucleoproteínas (RNP) devem ser transportadas ativamente através da CPN. Isso significa que a energia da hidrólise ATP e GTP é necessária. Cada molécula é transportada de uma maneira específica.
Em geral, as moléculas de RNA são embaladas com complexos RNP formadores de proteínas, sendo exportados dessa maneira. Por outro lado, as proteínas transportadas ativamente para o núcleo devem ter um sinal de sequência de localização no núcleo (SLN), possuidor de resíduos de aminoácidos com carga positiva (por exemplo, KKKRK).
Pode atendê -lo: mesosomeProteínas exportadas para o núcleo devem ter um sinal de exportação para o núcleo (NES) rico em aminoácido de leucina.
Além de facilitar o transporte entre o núcleo e o citoplasma, os CPNs estão envolvidos na organização da cromatina, regulação da expressão gênica e reparo de DNA. Nucleoporinas (NUPs) promovem a ativação ou repressão da transcrição, independentemente do estado de proliferação celular.
Em leveduras, as Nupts são encontradas no CNP do embrulho nuclear. Nos metazoários eles estão dentro. Eles desempenham as mesmas funções em todos os eucariotos.
Importação de substâncias
Através da CPN, há disseminação passiva de pequenas moléculas em direções e transporte ativo, importação de proteínas, exportação de RNA e ribonucleoproteínas (RNPs) e as moléculas bidirecionais Shuttle. O último inclui RNA, RNP e proteínas envolvidas na sinalização, biogênese e substituição.
A importação de proteínas dentro do núcleo ocorre em duas etapas: 1) ligação de proteínas ao lado citoplasmático do CPN; 2) translocalização dependente de ATP através do CPN. Este processo precisa de hidrólise ATP e a troca de GTP/PIB entre o núcleo e o citoplasma.
De acordo com um modelo de transporte, o complexo de proteínas receptores se move ao longo do canal através da união, dissociação e união novamente para as repetidas sequências FG das nucleoporinas. Dessa forma, o complexo se move de uma nucleoporina para outra dentro do CPN.
Exportação de substâncias
É semelhante à importação. RAN GTPase impõe direcionalidade no transporte através do CNP. RAN é um interruptor molecular com dois estados conformacionais, dependendo se está vinculado ao PIB ou GTP.
Duas proteínas reguladoras específicas desencadeiam a conversão entre os dois estados: 1) proteína ativadora da GTPase citosólica (GAP), que produz hidrólise de GTP e, portanto, converte RAN-GTP em RAN-GDP; e 2) fator de troca de guanina nuclear (GEF), que promove a troca de PIB pelo GTP e converte o RAN-GDP em RAN-GTP.
O citosol contém principalmente o RAN-GDP. O núcleo contém principalmente ran-gtp. Este gradiente das duas formas conformacionais de RAN direciona o transporte no endereço apropriado.
A importação do receptor, juntamente com a posição, é facilitada pela União para as repetições-FG. Se você chegar ao lado nuclear do CNP, o Ran-GTP se junta ao receptor para liberar sua posição. Assim, o RAN-GTP cria a direção do processo de importação.
A exportação nuclear é semelhante. No entanto, Ran-GTP no núcleo promove a união da posição para o receptor de exportação. Quando o receptor de exportação se move através do poro em direção ao citosol, ele encontra o Ran-gap, o que induz hidrólise GTP ao PIB. Finalmente, o receptor é liberado de sua posição e RAN-GDP no citosol.
Pode atendê -lo: leucócitos polimorfonuclearesTransporte de RNA
A exportação de alguns tipos de RNA é semelhante à exportação de proteínas. Por exemplo, Arnt e RNans (pequenos nucleares) usam o gradiente Rangtp e são transportados pelo CPN por meio. A exportação de ribossomos maduros também depende do gradiente Rangtp.
O mRNA é exportado de uma maneira muito diferente das proteínas e de outros RNAs. Para sua exportação, o mRNA forma um complexo RNP Messenger (RNPM), no qual uma molécula RNM é cercada por centenas de moléculas de proteína. Essas proteínas têm o processamento, Captura, Empalme e Polyadentilly of MNA.
A célula deve ser capaz de distinguir entre RNPM com RNM maduro e RNPM com mRNA imaturo. O mRNA, que forma o complexo RPNM, poderia adotar topologias que precisam ser remodeladas para transporte. Antes de entrar no RNPM no CPN, ocorre uma etapa de controle realizada por complexos de proteínas de Tramp e Exossomos.
Quando o RNPM maduro é montado, o RPNM é transportado pelo canal por meio de um receptor de transporte (NXF1-NXT1). Este receptor precisa de hidrólise de ATP (não um gradiente de Rangtp) para estabelecer a remodelação da direcionalidade do RNPM, que atingirá o citoplasma.
O complexo de poros nucleares e o controle da expressão gênica
Alguns estudos indicam que os componentes da CPN podem influenciar a regulação da expressão genética através do controle da estrutura da cromatina e sua acessibilidade a fatores de transcrição.
Nos eucariotos de evolução mais recente, a heterocromatina está preferencialmente localizada na periferia do núcleo. Este território é interrompido pelos canais de eucromatina, que são mantidos pela cesta nuclear da CPN. A associação de cesto nuclear com a euchromatina está relacionada à transcrição de genes.
Por exemplo, a ativação da transcrição no nível da CPN implica a interação dos componentes da cesta nuclear com proteínas como a saga da histona acetiltransferase e os fatores de exportação de RNA.
Assim, a cesta nuclear é uma plataforma para numerosos genes de manutenção (Serviço de limpeza) altamente transcritos e genes fortemente induzidos por mudanças nas condições ambientais.
O complexo de poros e virologia nuclear
A infecção por células virais eukaryot depende da CPN. Em cada caso de infecção viral, seu sucesso depende do DNA, RNA ou RPN passando pelo CPN para atingir seu objetivo final, que é a replicação do vírus.
Pode servir a você: inclusões citoplasmáticas: o que são, características, funçõesO vírus 4 40 (SV40) tem sido um dos modelos mais estudados para investigar a função do CPN na translocalização dentro do núcleo. Isso ocorre porque o SV40 tem um pequeno genoma (5.000 bases).
Foi demonstrado que o transporte do DNA do vírus é facilitado proteínas do vírus, que protegem o vírus até que o núcleo chegue.
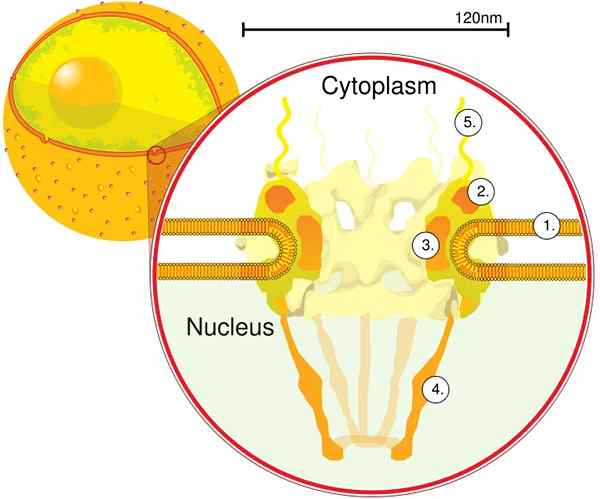
Componentes
O CPN é incorporado dentro do envelope nuclear e é composto entre aproximadamente 500 e 1000 notes. Essas proteínas são organizadas em subcomplexos ou módulos estruturais, que interagem entre si.
O primeiro módulo é um componente central, ou anel, dentro do poro na forma de um relógio de areia, que é limitado por outro anel de 120 nm de diâmetro em ambos os lados, intranucleares e citoplasmáticos. O segundo módulo é o núcleo e os anéis de citoplasma (120 nm de diâmetro cada) localizados ao redor do componente em forma de sandues.
O terceiro módulo são oito filamentos projetados a partir do anel de 120 nm dentro do nucleoplasma e forma uma estrutura em forma de cesta. O quarto módulo é composto pelos filamentos projetados ao lado do citoplasma.
O complexo em forma de y, composto por seis proteínas NUPs e SEH 1 e Sec 13, é o maior e mais caracterizado complexo do CNP. Este complexo é a unidade essencial que faz parte do andaime da CPN.
Apesar da baixa semelhança entre as seqüências NUPs, o andaime da CPN é altamente preservado em todos os eucariotos.
Referências
- Beck, m., Magoado, e. 2016. O complexo de poros nucleares: entender sua função através da percepção estrutural. Revisões da natureza, biologia celular molecular, doi: 10.1038/nrm.2016.147.
- Ibarra, a., Hetzer, m.C. 2015. Proteínas de poros nucleares e o controle das funções do genoma. Genes and Development, 29, 337-349.
- Kabachinski, g., Schwartz, t.OU. 2015. O complexo de poros nucleares - estrutura e função de relance. Journal of Cell Science, 128, 423-429.
- Knocknhauer, k.E., Schwartz, t.OU. 2016. O complexo de poros nucleares como portão flexível e dinâmico. Cell, 164, 1162-1171.
- Ptak, c., Aitchison, J.D., Wozniak, r.C. 2014. O nuclear multifuncional por complexo: uma plataforma para controlar a expressão gênica. Opinião atual da biologia celular, doi: 10.1016/j.CEB.2014.02.001.
- Stawicki, s.P., Steffen, J. 2017. Republicação: o complexo de poros nucleares - uma revisão abrangente da estrutura e função. International Journal of Academic Medicine, 3, 51-59.
- Tran, e.J., Goe, s.R. 2006. Nuclear dinâmico por complexos: vida no limite. Célula, 125, 1041-1053.