

Esfingolipídeos o que são, características, funções, síntese
- 3316
- 256
- Melvin Mueller
O que são esfidelipolipídios?
O esfingolipídios Eles representam uma das três grandes famílias lipídicas presentes nas membranas biológicas. Como glicofosfolipídios e esteróis, são moléculas anfípicas com uma região polar hidrofílica e uma região apolar hidrofóbica.
Eles foram descritos pela primeira vez em 1884 por Johann L. C. Thudichum, que descreveu três esfingolipídios (esfingomina, cérebro e cérebro) que pertencem às três classes diferentes que são conhecidas: fosfosfingolipídios, glicosfingolipídios neutros e ácidos.
Ao contrário dos glicofosfolipídios, os esfodyingolipids não são construídos em uma molécula de glicerol de 3 fosfato como o esqueleto principal, mas são compostos derivados do esfinição, um aminoalcohol com uma cadeia de hidrocarbonetos longa presa por uma ligação amida.
EEstrutura dos esfingolipídios
Todos os esfingolipídios derivam de uma serina L, que é condensada com um ácido graxo de cadeia longa para formar a base esfingóide, também conhecida como base de cadeia longa (LCB).
As bases mais comuns são a esfinganina e a esfinge, que diferem entre si na presença de uma ligação dupla Trans Entre carbonos 4 e 5 do ácido graxo da esfinge.
Os carbonos 1, 2 e 3 da esfinge são estruturalmente análogos aos glicerofosfolipídios de glicerol glicerol. Quando as ligações amida se ligam a um ácido graxo ao carbono 2 do Sphin.
Ácidos graxos de cadeia longa que constituem as regiões hidrofóbicas desses lipídios podem ser muito diversas. Os comprimentos variam de 14 a 22 átomos de carbono que podem ter diferentes graus de saturação, geralmente entre carbonos 4 e 5.
Nas posições 4 ou 6, eles podem ter grupos hidroxili e ligações duplas em outras posições ou mesmo ramificações, como grupos metila.
Caracteristicas
As cadeias de ácidos graxos ligados por ligações amida às ceramídeos são comumente saturadas e tendem a ter maior comprimento do que as encontradas nos glicerofosfolipídios, o que parece ser crucial para a atividade biológica destes.
Uma característica distinta do esqueleto de esfingolipídeo é que eles podem ter uma carga líquida positiva para pH neutro, estranho entre as moléculas lipídicas.
No entanto, o PkUm do grupo amino é baixo em relação a uma amina simples, entre 7 e 8, portanto, uma parte da molécula não é carregada com pH fisiológico, o que poderia explicar o movimento "livre" desses entre Bilapas.
A classificação tradicional de esfingolipídeos surge das múltiplas modificações que a molécula de ceramida pode sofrer, especialmente em termos das substituições dos grupos de cabeça polar.
Pode servir: hepadnavírus: características, morfologia, tratamentoFunções
Esfingolipídios são essenciais em animais, plantas e fungos, bem como em alguns organismos e vírus procarióticos.
-Funções estruturais
Os esfingolipídios modulam as propriedades físicas das membranas, incluindo sua fluidez, espessura e curvatura. Modular Essas propriedades também oferecem influência direta na organização espacial das proteínas da membrana.
Em "balsas" lipídicas
Nas membranas biológicas, os micro domínios dinâmicos podem ser detectados com menor fluência formada por colesterol e moléculas de esfingolipídeos chamadas lipídios balsas.
Essas estruturas ocorrem naturalmente e mantêm uma relação estreita com proteínas abrangentes, receptores de superfície celular e proteínas de sinalização, transportadores e outras proteínas com âncoras de glicosilfosfatidilinitol (GPI) (GPI).
-Funções de sinalização
Eles têm funções como moléculas de sinalização que atuam como segundos mensageiros ou como ligantes secretados para receptores de superfície celular.
Como mensageiros secundários, eles podem participar da regulamentação da homeostase do cálcio, crescimento celular, tumorogênese e supressão da apoptose. Além disso, a atividade de muitas proteínas de membrana integral e periférica depende de sua associação com esfingolipídeos.
Muitas interações intercelulares e celulares com seus arredores dependem da exposição dos diferentes grupos polares dos esfingolipídios em direção à face externa da membrana plasmática.
A união dos glicosfingolipídios e lectinas é crucial para a associação da mielina com axônios, a adesão de neutrófilos ao endotélio, etc.
Por -produtos de seu metabolismo
Os esfingolipídios de sinalização mais importantes são as bases de cadeia longa ou as esfingos e ceramídeos, bem como seus derivados fosforilados, como a esfigossina 1-fosfato.
Os produtos do metabolismo de muitos esfingolipídios ativam ou inibem múltiplos alunos brancos (proteínas quinase, fosfatose e outros), que controlam comportamentos celulares tão complexos quanto crescimento, diferenciação e apoptose.
-Como receptores de membrana
Alguns patógenos usam gloysphingolipids como receptores para mediar sua entrada para células hospedeiras ou para fornecer fatores de virulência a eles.
Foi demonstrado que os sfiyglipids participam de vários eventos celulares, como secreção, endocitose, quimiotáxia, neurotransmissão, angiogênese e inflamação.
Eles também estão envolvidos no tráfego de membrana, então influenciam a internalização de receptores, ordem, movimento e fusão de vesículas secretoras em resposta a diferentes estímulos.
Grupos esfingolipídicos
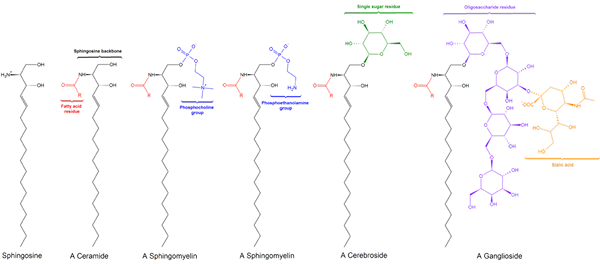
Existem três subclasses de esfingolipídeos, todas derivadas da ceramida e que diferem entre si por grupos polares, a saber: sfhilingomielinas, glicolipídios e gângliasides.
Esfingomyielinas
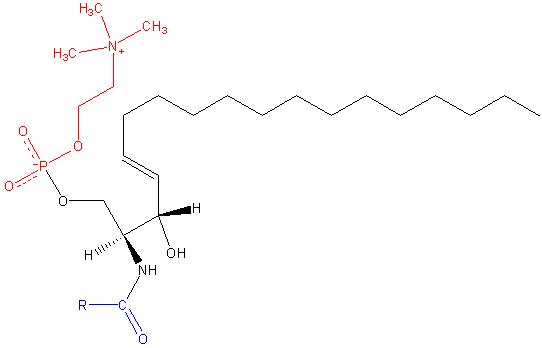 Sfingomilein. Black: Esfinge. Vermelho: fosfocolina. Azul: ácido graxo.
Sfingomilein. Black: Esfinge. Vermelho: fosfocolina. Azul: ácido graxo. Estes contêm fosfocolina ou fosfoetanolamina como um grupo de cabeça polar, para que sejam classificados como fosfolipídios junto com glicofosfolipídios. Eles olham, é claro, as fosfatidilcolinas na estrutura tridimensional e nas propriedades gerais, pois não têm carga em suas cabeças polares.
Pode atendê -lo: enolase: estrutura, mecanismo de ação, funçõesEles estão presentes nas membranas plasmáticas das células animais e são especialmente abundantes em mielina, uma vagem que circunda e isola os axônios de alguns neurônios.
Glicolipídios ou glicosfingolipídios neutros (sem carga)
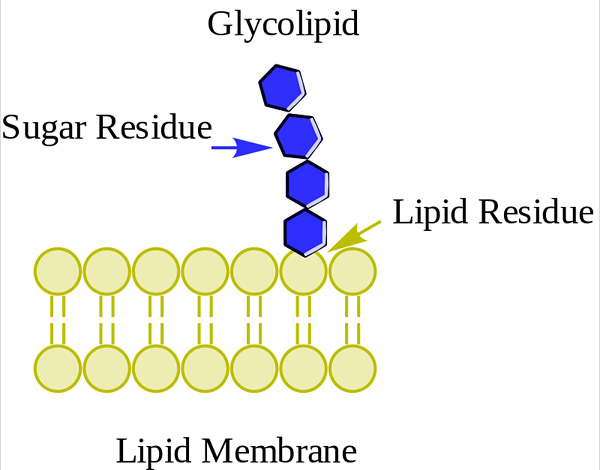 Glucolipídeo. WPCROSSON [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenças/BY-SA/4.0)], da Wikimedia Commons
Glucolipídeo. WPCROSSON [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenças/BY-SA/4.0)], da Wikimedia Commons Eles estão principalmente na face externa da membrana plasmática e têm um ou mais açúcares como um grupo de cabeça polar diretamente unida ao carbono hidroxil 1 da porção de ceramida. Eles não têm grupos de fosfato. Como o pH 7 não tem carga, eles são chamados de glicolipídios neutros.
Os cerebrosídeos têm uma única molécula de açúcar ligada à cerâmide. Aqueles que contêm galactose estão nas membranas plasmáticas de células de tecido não nervoso. Os globosídeos são goustertoesphingolipids com dois ou mais açúcares, normalmente D-glicose, D-galactose ou N-acetil-D-galactosamina.
Gangliósidos ou glicosfingolipídios
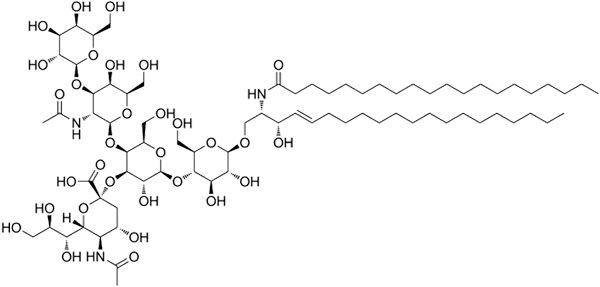 GM1 Estrutura ganglosida
GM1 Estrutura ganglosida Estes são os esfingolipídios mais complexos. Eles têm oligossacarídeos como um grupo de cabeça polar e um ou mais resíduos de ácido n-acetilmurâmico terminais, também chamados de ácido siral. O ácido siamico traz para os gânglios a carga negativa para pH 7, que os distingue de glicosfingolipídios neutros.
A nomenclatura desse tipo de esfingolipídeos depende da quantidade de resíduos de ácido sinalico presente na porção de oligossacarídeos da cabeça polar.
Síntese
A molécula de base longa ou esfinante é sintetizada no retículo endoplasmático (ER) e a adição do grupo polar à frente desses lipídios ocorre posteriormente no complexo de Golgi. Nos mamíferos, alguma síntese de esfingolipídios também pode ocorrer em mitocôndrias.
Depois de concluir sua síntese no complexo Golgi, os esfingolipídios são transportados para outros compartimentos celulares através de mecanismos mediados por vesículas.
A biossíntese dos esfingolipídios consiste em três eventos fundamentais: a síntese das bases de cadeia longa, a biossíntese das ceramidas pela união de um ácido graxo através de uma ligação amida e, finalmente, a formação dos complexos esfingolipídeos por meio da união de grupos polares no carbono 1 da base de sfhingoide.
Além da síntese de Novo, Os esfingolipídios também podem se formar pela substituição ou reciclagem das bases e ceramídios de cadeia longa, que podem alimentar a piscina esfingolipídica.
Síntese de esqueleto de ceramida
A biossíntese da ceramida, o esqueleto dos esfingolipídios, começa com a condensação descarboxillerativa de uma molécula de palmitail-coa e um L-Serine. A reação é catalisada por uma serina palmitail transferase (SPT) dependente de fosfato piridoxal e o produto é a di-hidroesfingosina ZE de 3 zo ze.
Pode atendê -lo: microbiota: composição, funções e onde estáEsta enzima é inibida por β-phaal-l-alaninas e l-cicloserinas. Em leveduras, é codificado por dois genes, enquanto em mamíferos existem três genes para esta enzima. O local ativo está na face citoplasmática do retículo endoplasmático.
O papel desta primeira enzima é preservado em todos os organismos estudados. No entanto, existem algumas diferenças entre os táxons que têm a ver com a localização subcelular da enzima: as bactérias são citoplasmáticas, leveduras, plantas e animais estão no retículo endoplasmático.
A 3-Zesfinganine é subsequentemente reduzida pela 3-zo-chetosfinganina dependente de Nadph. A acetila da di-hidroceramida sintase (esfinganina n-acil transferase) para produzir di-hidroceramida. A ceramida é então formada pela di -hidroceramida deaturase/redutase, que insere uma ligação dupla Trans Na posição 4-5.
Nos mamíferos, existem inúmeras isoformas de ceramida sintasas, cada uma se juntando a uma cadeia de ácidos graxos específicos às bases de cadeias longas. Portanto, as ceramidas synthesas e outras enzimas, elongasas, fornecem a principal fonte de diversidade de ácidos graxos em esfingolipídios.
Metabolismo
A degradação dos esfingolipídeos é realizada pelos glico -hidroles e enzimas esfingomielinase, responsáveis por remover as modificações dos grupos polares. Por outro lado, as ceramidases regeneram as bases de cadeia longa das ceramídios.
Os gânglios são degradados por um conjunto de enzimas lisossômicas que catalisam o passo a passo das unidades de açúcar, produzindo uma ceramida no final.
Outra maneira de degradação é a internalização de esfingolipídios nas vesículas endocíticas que são enviadas de volta à membrana plasmática ou transportadas para lisossomos, onde são degradadas por hidrolase ácida específica.
Nem todas as bases de cadeia longa são recicladas, a retenção endoplasmática. Esse mecanismo de degradação consiste em fosforilação em vez de acilação dos LCBs, dando origem a moléculas de sinalização que podem ser substratos solúveis para a enzima Liases.
Regulamento
O metabolismo desses lipídios é regulado em vários níveis, um deles é o das enzimas encarregadas da síntese, suas modificações pós -traducionais e os mesmos mecanismos do mesmo.
Alguns mecanismos de regulação são especificações celulares, para controlar o tempo do desenvolvimento de células em que são produzidos ou em resposta a sinais específicos.
Referências
- Bartke, n., & Hannun, e. (2009). Spphingolipids bioativos: metabolismo e função. Journal of Lipid Research, cinquenta, 19.
- Breslow, d. K. (2013). Homeostase esfingolipídica no retículo endoplasmático e além. Perspectivas frias de Spring Harbor em biologia, 5(4), A013326.