

Estrutura de uracil, funções, propriedades, síntese
- 2954
- 321
- Ralph Kohler
Ele Uracil É uma base de nitrogênio pirimidina, encontrada no ácido ribonucleico (RNA). Esta é uma das características que diferenciam o RNA do ácido desoxirribonucleico (DNA), já que este tem Timina em vez de Uracil. Ambas as substâncias, uracil e timina, diferem apenas que o segundo tem um grupo metil.
Do ponto de vista evolutivo, foi proposto que o RNA foi a primeira molécula que armazenou informações genéticas e trabalhou como catalisador nas células, antes de DNA e enzimas. Por causa disso, pensa -se que Uacil teve um papel fundamental na evolução da vida.
Fonte: Kemikungen [domínio público] Nos seres vivos, o uracil não é encontrado livremente, mas geralmente nucleotídeos de monofosfato (UMP), difosfato (UDP) e trfosfato (UTP). Esses nucleotídeos de uracil têm funções diferentes, como biossíntese de RNA e glicogênio, interconversão isomérica de açúcares e regulação da sintamina sintase.
[TOC]
Estrutura e propriedades
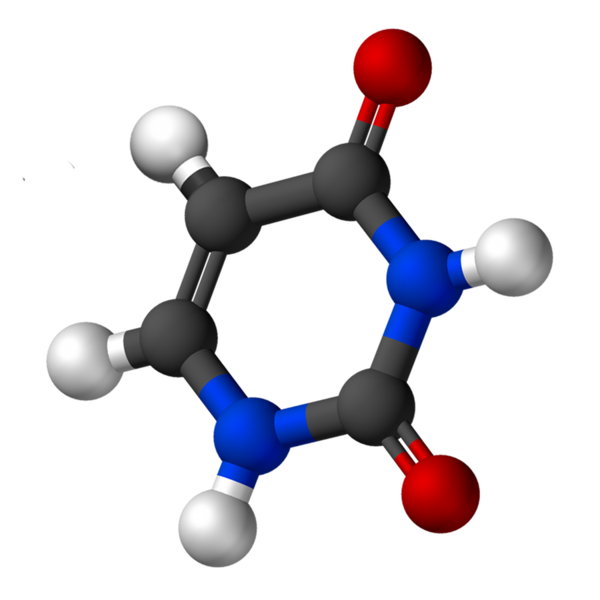
Uracil, chamado 2.4-dioxipiridina, tem a fórmula empírica C4H4N2QUALQUER2, cujo peso molecular é 112,09 g/mol e é purificado como um pó branco.
A estrutura da uridina é um anel heterocíclico com quatro átomos de carbono e dois nitrogênio, com ligações alternativas duplas. É planar.
Possui uma solubilidade de 50 mg/ml, 25 ºC, em 1m de hidróxido de sódio e um PKA entre 7.9 e 8.2. O comprimento de onda onde ocorre sua absorvância máxima (ʎMáx) Está entre 258 e 260 nm.
Biossíntese
Existe um caminho comum para a biossíntese de nucleotídeos de pirimidina (uracil e citocina). O primeiro passo é a biossíntese de fosfato de carbamoil de CO2 e NH4+, que é catalisado por fosfato de carbamoil sintetase.
A pirimidina é construída a partir de fosfato de carboil. Ambas as substâncias reagem e formam n-carbamoilaspartato, catalisado pelo Aspartato de Transcabamoilasa (ATCASA). O fechamento do anel de pirimidina pode ser desidratado pela di-hidrootase e produz l-di-hidrotatoato.
O L-di-hidrotoato é oxidado e transformado em orotato; O aceitador de elétrons é o NAD+. É uma reação catalisada por di -hidroorotato desidrogenase. A próxima etapa consiste na transferência do grupo fosforibosil, do fosforbosil pirofosfato (PRPP), para a orotate. Forma gradidilato (WPO) e piroffosfato inorgânico (PPI), catalisado pela fosforibosil orotate transferase.
O último passo consiste em descarboxilação do anel de pirimidina do grupo (WPO). Forma uridilaato (uridina-5'-monofosfato, ump), que é catalisada por uma descarboxilase.
Então, através da participação de uma quinase, um grupo de fosfato é transferido do ATP para o UMP, formando UDP (uridina-5'-difosfato). O último é repetido, formando UTP (uridina-5'-trifosfato).
Pode atendê -lo: ramos da biologia e o que eles estudamRegulação da biossíntese
Nas bactérias, a regulação da biossíntese de pirimidina ocorre através de feedback negativo, no nível de aspartato de transcabamoilasa (ATCASA).
Esta enzima é inibida pelo CTP (Citidin-5'-trofosfato), que é o produto final da via de biossíntese das pirimidinas. ATCAS.
Nos animais, a regulação da biossíntese de pirimidina ocorre através de feedback negativo, no nível de duas enzimas: 1) fosfato da carbamoil sintase II, que é inibido pelo UTP e ativado por ATP e PRPP; e 2) a OMP Discarboxilase, que é inibida pelo produto da reação que ela mesma catalisa, o UMP. A velocidade de biossíntese do OMP varia com a disponibilidade de prpp.
Função na biossíntese do RNA
O uracil está presente em todos os tipos de RNA, como o RNA mensageiro (MNA), o RNA de transferência (ARNT) e o RNA ribossômico (RNA). A biossíntese dessas moléculas ocorre através de um processo chamado transcrição.
Durante a transcrição, as informações contidas no DNA são copiadas no RNA através de um RNA de Pon Polimerase. O processo reverso, no qual as informações contidas no RNA são copiadas no DNA, acontece em alguns vírus e plantas através da transcriptase reversa.
A biossíntese de RNA precisa de nucleosídeos de triposfato (NTP), a saber: trifosfato uridina (UTP), citidina de triposfato (CTP), adenina adenina adenina (ATP) e trifosfato guanina (GTP). A reação é:
(RNA)n desperdício + Ntp -> (RNA)N+1 Resíduos + PPI
A hidrólise inorgânica de pirofosfato (PPI) fornece energia para a biossíntese de RNA.
Função na biossíntese de açúcar
Ésteres de açúcar são muito comuns em organismos vivos. Alguns desses ésteres são os difosfas dos ésteres de nucleosídeos, como UDP-Sugar, que são muito abundantes nas células. O UDP-Sugar envolve a biossíntese de dissacarídeos, oligossacarídeos e polissacarídeos.
Nas plantas, a biossíntese de sacarose acontece de duas maneiras: um principal e um secundário.
A rota principal consiste na transferência da D-glicose do UDP-D-Glucosa para o D-Frucease para formar sacarose e UDP. A estrada secundária inclui duas etapas: começa com UDP-D-glicose e frutose-6-fosfato e culmina com a formação de sacarose e fosfato.
Nas glândulas mamárias, a biossíntese de lactose ocorre a partir de udp-d-galactose e glicose.
Pode atendê -lo: ramos da bioquímicaNas plantas, a biossíntese de celulose é realizada por condensação contínua de resíduos beta-d-glicosil, da UDP-glicose à extremidade não redutora da cadeia de policlucose em crescimento. Da mesma forma, a biossíntese de amilose e amilopetina requer UDP-glicose como substrato doador de glicose para a cadeia de crescimento.
Em animais, tanto o UDP-glicose quanto o ADP-glicose são usados para a biossíntese de glicogênio. Da mesma forma, a biossíntese de sulfato de condroitina requer udp-xilose, udp-galactose e udp-glucuronato.
Função na interconversão isomérica de açúcares
A conversão da galactose em um intermediário de glicólise acontece através da estrada Leloir. Uma das etapas desta rota é catalisada pela enzima UDP-Galactose-4-epimerase, que facilita a interconversão da UDP-Galactose para UDP-Glucose.
Função na biossíntese de glicoproteínas
Durante a biossíntese de glicoproteínas, as proteínas passam pelos sacos cis, médios e trans do aparelho de Golgi.
Cada uma dessas sacolas possui um conjunto de enzimas que processam glicoproteínas. Monômeros de açúcar, como glicose e galactose.
Os nucleotides-hexose são transportados para tanques de Golgi por AntiPorte. O UDP-Galactose (UDP-GAL) e o UDP-N-acetilgalactosamina (UDP-Galnac) entram nos tanques do citosol por meio de troca pela UMP.
No tanque de Golgi, uma fosfatase hidrolisa um grupo de fosfato da forma UDP e UMP e Pi. O UDP vem das reações catalisadas pela galactosiltransferase e N-acetilgalactosamiltransferase. O UMP formado pela fosfatase serve para trocar nucleotídeos-hexose.
Função na regulação da sintase
Um mecanismo para regular a glutamina sintase é a modificação covalente, que consiste em adenilação, que inativa e flanco, que a ativa. Esta modificação covalente é reversível e catalisada por adeniltransferase.
A atividade da adeniltransferase é modulada pela união da proteína PII, que é regulada por uma modificação covalente, uridinilação.
Tanto a uridililação quanto o deslocamento são realizados pela uridililtransferase. Nesta enzima, a atividade de uridilação é devida à glutamina e fosfato e é ativada pela união de alfa-zotoglutarato e ATP para o PII.
Função na edição de RNA
Alguns RNM são editados antes da tradução. Em alguns organismos eucarióticos, como Trypanosoma brucei, Há edição do RNA da transcrição do gene da subunidade II da citocromo oxidase. Isso acontece inserindo resíduos de uracil, uma reação catalisada pela uridiltransferase terminal.
Pode atendê -lo: cacho: composição, peças, importânciaUm RNA -guia, complementar ao produto editado, atua como temperado para o processo de edição. Os pares de bases formados entre a transcrição inicial e o RNA do guia implica pares de bases g = u que não são Watson-Record e são comuns no RNA.
Biossíntese de UDP-glicose
Em condições fisiológicas, a biossíntese de glicogênio da glicose-1-fosfato é termodinamicamente impossível (ΔG positivo). Por causa disso, antes da biossíntese, a ativação do glicose-1-fosfato (G1P) acontece. Este G1P e UTP combina reação para formar uridina de difosfato de glicose (UDP-glicose ou UDPG).
A reação é catalisada pela pirofosforlase da UDP-glicose e é a seguinte:
G1p + utp -> udp -glucosa + 2pi.
A variação de energia livre de Gibbs nesta etapa é grande e negativa (-33,5 kJ/mol). Durante a reação de oxigênio, o G1P ataca o átomo de fósforo alforo alfa da UDP-glicose e pirofosfato inorgânico (PPI). Então, o PPI é hidrolisado por um pirofosfato inorgânico, cuja energia de hidrólise é a que impulsiona a reação geral.
UDP-glicose é uma substância de "alta energia". Permite formar as ligações glicosídicas entre o resíduo de glicose e a crescente cadeia de polissacarídeos. Esse mesmo princípio de energia é aplicável às reações em que o udp-açúcar participa, como a biossíntese de dissacarídeos, oligossacarídeos e glicoproteínas.
Uracil DNA glicosilasa
Existem lesões de DNA que ocorrem espontaneamente. Uma dessas lesões é o sprontono. Nesse caso, o reparo ocorre devido à base de DNA modificada por meio de uma enzima chamada uracil DNA glicosilasa.
A enzima uracil DNA glicosilasa elimina a citocina danificada (uracil), produzindo um resíduo de desoxirribose que não possui a base de nitrogênio, chamada local de AP (local apurinico-apirimidínico).
Então, a enzima AP endonuclease faz um corte no esqueleto de fosfodiester do local da AP, eliminando o resíduo de açúcar-fosfato. DNA polimerase I restaura o fio danificado.
Referências
- Bohinski, r. 1991. Bioquímica. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Bioquímica. Editorial Reverté, Barcelona.
- Lodish, h., Berk, a., Zipurski, s.eu., Matsudaria, p., Baltimore, d., Darnell, J. 2003. Biologia celular e molecular. Medica editorial Panamericana, Buenos Aires, Bogotá, Caracas, Madri, México, Sāo Paulo.
- Nelson, d. eu., Cox, m. M. 2008. Lehninger-princípios da bioquímica. C.H. Freeman, Nova York.
- Voet, d. e Voet, J. 2004. Bioquímica. John Wiley e Sons, EUA.
- « Fórmula de custo marginal, cálculo e exercícios
- Características de pirimidinas, estrutura, funções »