

Estrutura da citosina, funções, propriedades, síntese
- 1181
- 247
- Conrad Schmidt
O Citosina É uma base de nitrogênio pirimidina, que serve para a biossíntese de citidina-5'-monofosfato e dexicicitidina-5'-monofosfato. Esses compostos servem para a biossíntese, respectivamente, do ácido desoxirribonucleico (DNA) e ácido ribonucleico (RNA). O DNA armazena informações genéticas e RNA tem várias funções.
Nos seres vivos, a citosina não é encontrada livremente, mas geralmente ribonucleotídeos ou desoxirribonucleotídeos. Ambos os tipos de composto têm um grupo de fosfato, uma base de ribose e nitrogênio.
Fonte: Vesprcom [Domínio Público] O carbono 2 da ribose possui um grupo oxidrillo (-OH) nos ribonucleotídeos e um átomo de hidrogênio (-h) no desoxirribonucleotídeo. Dependendo do número de grupos de fosfato presentes, existe citidín-5'-monofosfato (CMP), Citidín-5'-diposfato (CDP) e Citidín-5'-trifosfato (CTP) (CTP).
Os equivalentes desoxigenados são chamados desoxicitidina-5'-monofosfato (DCMP), DeSoxicicitidin-5'-diposfato (DCDP) e DeSoxicicitidin-5'-trifosfato (DCTP).
A citosina, em suas várias formas, participa de diferentes funções, como a biossíntese de DNA e RNA, a biossíntese de glicoproteínas e a regulação da expressão do gene.
[TOC]
Estrutura e propriedades
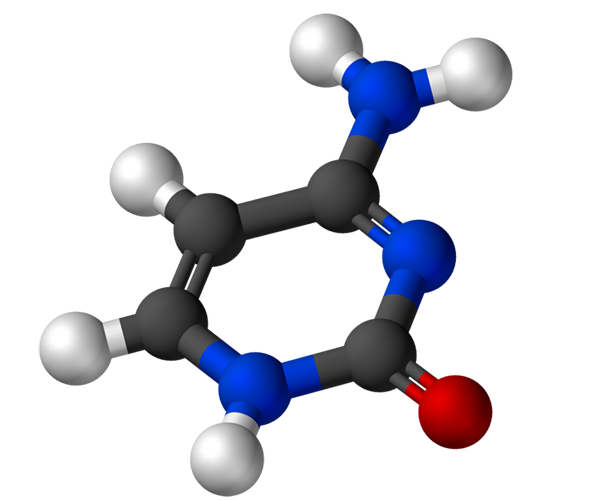
A citosina, 4-amino-2-hidroxipirimidina, tem a fórmula empírica C4H5N3Ou, cujo peso molecular é 111,10 g/mol e é purificado como um pó branco.
A estrutura da citosina é um anel heterocíclico, aromático e planar. O comprimento de onda máxima de absorvância (ʎMáx) É 260 nm. A temperatura de fusão da citosina excede 300 ºC.
Para formar um nucleotídeo, a citosina se liga covalentemente, através do nitrogênio 1, por meio. O carbono 5 'é esterificado com um grupo de fosfato.
Biossíntese
A biossíntese de nucleotídeos de pirimidina tem uma via comum, que consiste em seis etapas catalisadas por enzimas. A estrada começa com a biossíntese de carbamoil-fosfato. Nos procariontes, existe apenas uma enzima: fosfato carbamoil sintase. Isso é responsável pela síntese de pirimidinas e glutamina. Nos eucariotos, existem fosfato de carbamoil sintase I e II, que são responsáveis, respectivamente, pela biossíntese de glutamina e pirimidina.
O segundo passo consiste na formação de n-carbamoilaspartato, de fosfato de carboils e aspartato, reação catalisada pelo transpartato transcabamoilasa (atcasa).
O terceiro passo é a síntese de L-di-hidrotoato, que produz o fechamento do anel de pirimidina. Esta etapa é catalisada por di -hidrootase.
O quarto passo é a formação de Orotato, que é uma reação redox catalisada por di -hidroorotato desidrogenase.
A quinta etapa consiste na formação de ouro (OMP) usando fosforibosil pirofosfato (PRPP) como substrato e a fosforibossil orotato transferase como catalisador como catalisador.
O sexto passo é a formação de uridilaato (uridina-5'-monofosfato, UMP), catalisada por uma omp-decarboxilase.
Pode atendê -lo: flora e fauna de HidalgoAs etapas a seguir consistem na fosforilação da UMP, catalisada por cinases, para formar UTP e a transferência de um grupo amino de glutamina para o UTP para formar CTP, reação catalisada pela CTP sintetase.
Regulação da biossíntese
Nos mamíferos, a regulamentação ocorre na fosfato carbamoil sintase.
Carbamoil Syntasa II é regulamentado por feedback negativo. Seus reguladores, o UTP e o PRPP, são, respectivamente, inibidor e ativador desta enzima.
Nos tecidos que não são hepáticos, a fosfato de carbamoil sintase II é a única fonte de fosfato de carbamoil. Enquanto estava no fígado, sob condições de excesso de amônia, a fosfato de carbamoil sintase I produz, nas mitocôndrias, o fosfato de carbamoil, que é transportado para o citosol, de onde entra no caminho da biossíntese de pirimidinas.
Outro ponto de regulamentação é a OMP-Discarboxilase, que é regulada pela inibição competitiva. O produto de sua reação, UMP, compete com o OMP para o local de ligação no OMP-Descarboxilasa.
Pirimidinas, como a citosina, são recicladas
A reciclagem de pirimidinas tem como uma função a reutilização de pirimidinas sem a necessidade de biossíntese de Novo e evite o caminho degradativo. A reação de reciclagem é catalisada pela fosforibosiltransferase pirimidina. A reação geral é a seguinte:
Pirimidina + prpp -> nucleosídeo pirimidina 5 '-monofosfato + ppi
Nos vertebrados, a fosforibosiltransferase pirimimidina é encontrada em eritrócitos. O substrato das pirimidinas desta enzima são uracil, timina e orotato. A citosina é indiretamente reciclada da uridina-5'-monofosfato.
Função na biossíntese de DNA
Durante a replicação do DNA, as informações contidas no DNA são copiadas no DNA através de uma polimerase de DNA.
A biossíntese de RNA precisa de desoxinucleotídeos trifosfato (DNTP), a saber: dexitimida trifosfato (DTTP), trifosfato de dexicitidia (DCTP), des -alxizidenina trifyse (DATP) e dexiguanina Tryfosfato (DGTP). A reação é:
(DNA)n desperdício + Dntp -> (DNA)N+1 Resíduos + PPI
A hidrólise inorgânica de pirofosfato (PPI) fornece energia para a biossíntese de RNA.
Função na estabilização da estrutura do DNA
No duplo DNUS Helix, uma purina, de uma corrente, está ligada à pirimidina, da cadeia oposta, por ligações de hidrogênio. Assim, a citosina está sempre ligada à guanina por três ligações de hidrogênio: a adenina está ligada a Timin por duas ligações de hidrogênio.
Pode atendê -lo: importância da biologiaAs ligações de hidrogênio são quebradas quando uma solução de DNA nativo purificada a pH 7 é submetida a temperaturas maiores que 80 ºC. Isso faz com que a hélice dupla de DNA forme duas cadeias separadas. Este processo é conhecido como desnaturação.
A temperatura na qual 50% do DNA é desnaturada é conhecida como temperatura de fusão (TM). Moléculas de DNA cuja guanina e citosina.
O exposto acima é o teste experimental de que um número maior de ligações de hidrogênio estabiliza melhor as moléculas de DNA nativas.
Função de regiões ricas em citosina no DNA
Recentemente, verificou -se que o DNA do núcleo celular humano pode adotar estruturas de motivos intercalados (IM). Essas estruturas são produzidas em regiões de citosina.
A estrutura IM consiste em quatro cadeias de DNA, ao contrário do clássico DNA de hélice dupla que possui duas correntes. Mais especificamente, duas cadeias duplex paralelas são intercaladas em uma orientação antiparalla e são mantidas juntas por um par de citosinas hemiprotonas (C: C: C+).
No genoma humano, as estruturas de IM são encontradas em regiões como promotores e telômeros. O número de estruturas de IM é maior durante a fase G1/S do ciclo celular, no qual a transcrição é alta. Essas regiões são locais de reconhecimento de proteínas envolvidos na ativação de máquinas transcricionais.
Por outro lado, nas regiões ricas em bases de guanina (c) consecutivas, o DNA tende a adotar a hélice forma A, em condições de desidratação. Este formulário é típico de RNA e bandas mistas de DNA-DNA duplo durante a transcrição e replicação, e em determinados momentos em que o DNA está ligado à proteína.
Foi demonstrado que as regiões com bases consecutivas de citosina criam um patch eletropositivo na fenda principal do DNA. Portanto, acredita -se que essas regiões se ligam às proteínas, que predispõem certas regiões genômicas de fragilidade genética.
Função na biossíntese de RNA
Durante a transcrição, as informações contidas no DNA são copiadas no RNA através de um RNA de Pon Polimerase. A biossíntese de RNA precisa de nucleosídeos de triposfato (NTP), a saber: Tryphyse citidina (CTP), triposfato uridina (UTP), adenina triposfato (ATP) e trifosfato guanina (GTP). A reação é:
Pode atendê -lo: Flora e Fauna de Campeche: Espécies Representativas(RNA)n desperdício + Ntp -> (RNA)N+1 Resíduos + PPI
A hidrólise inorgânica de pirofosfato (PPI) fornece energia para a biossíntese de RNA.
Função na biossíntese de glicoproteínas
A transferência seqüencial de hexoses para formar os oligossacarídeos, O-ligados para proteínas, ocorre de precursores de nucleotídeos.
Nos vertebrados, a última etapa da biossíntese de oligossacarídeos ligados a O consiste na adição de dois resíduos de ácido sinal (n-acetilneuramina) de um precursor de citidina-5'-monofosfato (CMP). Esta reação é produzida no saco trans de Golgi.
Tratamentos de citosina e quimioterapia contra o câncer
O ácido tetra -hidrofolato (FH4) é uma fonte de grupo -CH3, E é necessário para a biossíntese do DTMP do despejo. FH2 também é formado. A redução de FH2 para FH4 requer um folato e Nadph redutase. Alguns inibidores de folato redutase, como aminopterina e metretotrexato, são usados em tratamentos contra o câncer.
Metotrexan é um inibidor competitivo. O folato redutase se une a 100 vezes mais afinidade a esse inibidor do que ao seu substrato. A aminapterina atua da mesma forma.
A inibição da folato redutase dificulta indiretamente a biossíntese DTMP e, portanto, a do DCTP. A inibição direta ocorre através dos inibidores da timidilato sintetase, que catalisa o DTMP do despejo. Esses inibidores são 5-fluorouracil e o 5-fluoro-2-dioxiuridina.
Por exemplo, o 5-fluoroacil não é um inibidor, mas primeiro se torna, a rota de reciclagem, no mofosfato de desoxiurídeo D (FDUMP), que se liga à sintetase do timidilato e o inibe.
Substâncias análogas à glutamina, azaserina e acivicina, inibem a amidotransferase glutamina. Azarine foi uma das primeiras substâncias descobertas que atuam como inativação suicida.
Referências
- Assi, h.PARA., Garavís, m., González, c., E Damha, M.J. 2018. DNA I-Motif: características estruturais e significado para biologia celular. Nuclei Acids Research, 46: 8038-8056.
- Bohinski, r. 1991. Bioquímica. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Bioquímica. Editorial Reverté, Barcelona.
- Lodish, h., Berk, a., Zipurski, s.eu., Matsudaria, p., Baltimore, d., Darnell, J. 2003. Biologia celular e molecular. Medica editorial Panamericana, Buenos Aires, Bogotá, Caracas, Madri, México, Sāo Paulo.
- Nelson, d. eu., Cox, m. M. 2008. Lehninger-princípios da bioquímica. C.H. Freeman, Nova York.
- Voet, d. e Voet, J. 2004. Bioquímica. John Wiley e Sons, EUA.