

Síntese de tipos lipídicos e seus principais mecanismos
- 2958
- 290
- Tim Mann
O Síntese lipídica Consiste em uma série de reações enzimáticas por meio de quais hidrocarbonetos de cadeia curta são condensados para formar moléculas de cadeia mais longa que podem subsequentemente sofrer diferentes modificações químicas.
Os lipídios são um tipo muito variado de biomoléculas sintetizados por todas as células vivas e são especializadas em múltiplas funções essenciais para a manutenção da vida celular.
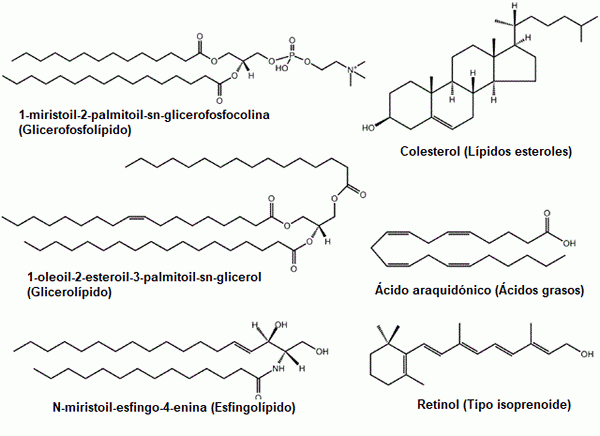 Alguns exemplos de lipídios comuns: glicerofosfolipídios, esteróis, glicerolipídios, ácidos graxos, esfingolipídios e pré -nóis (fonte: o uploader original era LMAPS na Wikipedia em inglês. / Gfdl 1.2 (http: // www.gnu.Org/licenças/licendas antigas/fdl-1.2.HTML) via Commons, adaptado por Raquel Parada)
Alguns exemplos de lipídios comuns: glicerofosfolipídios, esteróis, glicerolipídios, ácidos graxos, esfingolipídios e pré -nóis (fonte: o uploader original era LMAPS na Wikipedia em inglês. / Gfdl 1.2 (http: // www.gnu.Org/licenças/licendas antigas/fdl-1.2.HTML) via Commons, adaptado por Raquel Parada) Os lipídios são os principais componentes das membranas biológicas, um fato que os torna moléculas fundamentais para a existência de células como entidades isoladas de seus arredores.
Alguns lipídios também possuem funções especializadas, como pigmentos, cofatores, transportadores, detergentes, hormônios, mensageiros intra-extracelulares, âncoras covalentes para proteínas da membrana, etc. Portanto, a capacidade de sintetizar diferentes tipos de lipídios é fundamental para a sobrevivência de todos os organismos vivos.
Esse grande grupo de compostos é tradicionalmente classificado em várias categorias ou subgrupos: ácidos graxos (saturados e insaturados), lipídios complexos de glicerídeos (lipoproteínas).
[TOC]
Tipos de lipídios e seus principais mecanismos de síntese
Todas as seqüências de reação das rotas de biossíntese lipídica são endogônicas e redutivas. Em outras palavras, todos eles usam ATP como fonte de energia e um transportador de elétrons reduzido, como o NADPH, como um poder redutor.
Em seguida, serão descritas as principais reações das rotas biossintéticas dos principais tipos de lipídios, ou seja, de ácidos graxos e eicosanóides, triacilgliceroles e fosfolipídios e esteróis (colesterol) (colesterol).
- Síntese de ácidos graxos
Os ácidos graxos são moléculas extremamente importantes do ponto de vista lipídico, pois fazem parte dos lipídios mais relevantes nas células. Sua síntese, ao contrário do que muitos cientistas pensavam durante os primeiros estudos, não consistem na rota reversa de sua oxidação β.
De fato, essa rota metabólica ocorre em diferentes compartimentos celulares e requer a participação de um intermediário de três átomos de carbono conhecidos como malonil-CoA, o que não é necessário na oxidação.
 Malonil-coa. Domínio neuroteker / pub
Malonil-coa. Domínio neuroteker / pub Além disso, está intimamente relacionado aos grupos de proteínas de proteínas em sulfidilos conhecidas como transportadores de grupo acil (ACP, inglês Proteínas transportadoras de acila).
Em geral, a síntese de ácidos graxos, especialmente a da cadeia longa, é um processo seqüencial em que quatro etapas são repetidas em cada "retorno" e, durante cada retorno, há um grupo ácido saturado que é o substrato para o próximo, que implica outra condensação com uma nova molécula de malonil-coa.
Em cada turno ou ciclo de reação, a cadeia de ácidos graxos se estende por dois carbonos, até que um comprimento de 16 átomos (palmitato) atinja, após o que o ciclo sai.
Formação Malononil-CoA
Esse intermediário de três átomos de carbono é irreversivelmente formado a partir de acetil-CoA, graças à ação de uma enzima acetil-CoA carboxilase, que possui um grupo de próstata de biotina que está covalentemente ligado à enzima e que participa dessa catálise nesta catálise em duas etapas.
Nesta reação, um grupo carboxil derivado de uma molécula de bicarbonato (HCO3-) é transferido para a biotina em uma forma dependente de ATP, onde o grupo biotinil cumpre a função do "transportador temporário" da molécula enquanto transferências para acetil-CoA, produzindo malonil-coa.
Pode atendê -lo: teste de oxidase: fundação, procedimento e usosNa sequência de síntese de ácidos graxos, o agente redutor usado é o NADPH e os grupos ativadores são dois grupos Tiol (-SH) que fazem parte de um complexo multienzimático chamado ácido graxo sintase, que é o mais importante na catálise sintética sintética.
Nos vertebrados, o complexo de ácidos graxos sintase faz parte de uma única cadeia polipeptídica grande, na qual as 7 atividades enzimáticas características da rota de síntese são representadas, bem como a atividade hidrolítica necessária para liberar intermediários no final da síntese.
 Estrutura enzimática de ácidos graxos Syntasa (fonte: Boehringer Ingelheim/CC BY-SA (https: // criativeCommons.Org/licenças/BY-SA/4.0) via Wikimedia Commons)
Estrutura enzimática de ácidos graxos Syntasa (fonte: Boehringer Ingelheim/CC BY-SA (https: // criativeCommons.Org/licenças/BY-SA/4.0) via Wikimedia Commons) As 7 atividades enzimáticas deste complexo são: proteína de transporte do grupo acilo (ACP), acetil-CoA-ACP transacetilas cetoacil-acp redutase (KR), β-hidroxiacil-ACP dehyddratasa (HD) e raiva-ace respiratória (ER).
Antes que as reações de condensação possam ocorrer para montar a cadeia de ácidos graxos, os dois grupos Tiol no complexo enzimático são "carregados" com os grupos acil: antes de tudo, um acetil -COA é transferido para o grupo -SH de uma cisteína Na β-ceticoacil-ACP sintase do complexo, a reação catalisada pela enzima transacetila acetil-CoA-ACP (AT) (AT) (AT).
Posteriormente, um grupo malônico é transferido de uma molécula de malonil -coa para o grupo -Sh Group of the ACILO Group (ACP), faz parte do complexo de ácidos graxos sintase.
A sequência de quatro reações para cada "retorno" do ciclo de reação é a seguinte:
- Condensação: os grupos acetil e malonil "carregados" na enzima são condensados para formar uma molécula de acetoacetil -ACP, que está ligada à parte ACP através de um grupo -Sh. Nesta etapa, há uma molécula de CO2 e é catalisada pela β-zoacil-ACP sintase (o grupo acetil ocupa a posição de "metil terminal" do complexo acetoacetil-ACP).
- Redução do grupo carbonil: o grupo carbonil na posição C3 do acetoacetil-ACP é reduzido para formar D-β-hidroxibutiril-ACP, catalisado pela reação catalisada pelo β----quelacl-ACP redutase, que NADPH usa como doador eletrônico.
- Desidratação: os carbonos C2 e C3 dos D-β-hidroxibutiril-ACP são desprovidos de moléculas de água, formando uma ligação dupla que termina com a produção do novo composto Trans-∆2-butenoil-ACP. Este processo é mediado por uma enzima β-hidroxiacil-ACP desidratase (HD) (HD).
- Redução de ligação dupla: A ligação dupla do composto formada na etapa de desidratação é saturada (reduzida) para dar origem ao butiril-ACP pela reação catalisada pelo enzima raiva-acp redutase (ER), que também usa NADPH como agente redutor.
As reações de síntese ocorrem até que uma molécula de palmitato (16 átomos de carbono) seja formada, que é hidrolisada do complexo enzimático e liberada como um possível precursor para ácidos graxos de cadeias de comprimento maior, que são produzidas por sistemas de alongamento de ácidos graxos localizados no liso parte do retículo endoplasmático e nas mitocôndrias.
Pode atendê -lo: fauna e flora do mar peruanoAs outras modificações que essas moléculas podem sofrer, como as deaturas, por exemplo, são catalisadas por diferentes enzimas, que geralmente ocorrem no retículo endoplasmático liso.
- Síntese de eicosanóides
Os eicosanóides são lipídios celulares que têm funções como moléculas mensageiras de "curta duração", produzidas por alguns tecidos para se comunicar com seus tecidos vizinhos. Essas moléculas são sintetizadas a partir de ácidos graxos poliinsaturados de 20 átomos de carbono.
Prostaglandins
Em resposta a um estímulo hormonal, a enzima fosfolipase ataca os fosfolipídios da membrana e libera araquidonato do glicerol carbono 2. Este composto é convertido em prostaglandinas graças a uma enzima do retículo endoplasmático suave com atividade bifuncional: ciclooxigenase (COX) ou prostaglandina h2 sintase.
Thromboxan
As prostaglandinas podem ser convertidas em tromboxanos graças à Thromboxan sintase nas plaquetas do sangue (trombócitos). Essas moléculas participam das etapas iniciais da coagulação do sangue.
- Síntese de triacilgliceroles
Os ácidos graxos são moléculas fundamentais para a síntese de outros compostos mais complexos em células, como triacilgliceroles ou lipídios lipídicos glicerofosfolipídios (processos que dependem das necessidades metabólicas celulares).
Os animais produzem triacilgliceroles e glicofosfalipídeos de dois precursores comuns: o acil de gordura e 3-fosfato L-glicerol. A acila de gordura-coa é produzida por síntesas acil-coa que participam da β-oxidação, enquanto o 3-fosfato L-glicerol é obtido da glicólise e pela ação de duas enzimas alternativas: 3-fosfato glicerol desidrogenase e quinase glicerol.
Os triacilgliceroles são formados pela reação entre duas moléculas de Flo-CoA ACIL e uma molécula de diacilglicerol de 3 fosfato; Essas reações de transferência são catalisadas por transferases específicas.
Nesta reação, existe, inicialmente, o ácido fosfatídico, que é desfosforilado por uma fosfatase de fosfatídeo enzimática para produzir 1.2-diacilglicerol, que é capaz, novamente, de aceitar uma terceira molécula de gordura-coa acil, produzindo triiailclorlos.
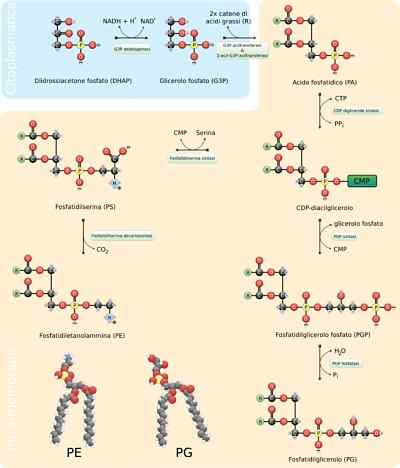
- Síntese de fosfolipídios
Os fosfolipídios são moléculas extremamente variáveis, como muitas diferentes podem se formar pela combinação de ácidos graxos e grupos "cabeça" com o esqueleto de glicerol (glicofosfolipídios) ou esfingosina (esfingolipídios) que os caracterizam.
A Assembléia Geral dessas moléculas requer a síntese do esqueleto de glicerol ou esférico, da união com os ácidos graxos correspondentes, seja por esterificação ou amidação, a adição de um grupo hidrofílico "cabeça" através de uma ligação fosfodiéster e, se necessário , a alteração ou troca desses últimos grupos.
Nos eucariotos, esse processo ocorre no retículo endoplasmático suave e também na membrana mitocondrial interna, onde podem permanecer indefinidamente ou de onde podem ser translocados para outros lugares.
Etapas de reação
As primeiras etapas da reação de síntese dos glicerofosfolipídios são equivalentes à produção de triacilgliceroles, pois uma molécula de glicerol de 3-fosfato é esterificada a duas moléculas de ácidos graxos em carbonos 1 e 2, formando fosfatídicos de ácido. É comum encontrar fosfolipídios com ácidos graxos saturados no C1 e insaturados no C2 de glicerol.
Pode atendê -lo: resistinaO ácido fosfatídico também pode ser produzido pela fosforilação de uma molécula de diacilglicerol já sintetizada ou "reciclada".
Os grupos polares "cabeça" dessas moléculas são formados através de links de fosfodiéster. A primeira coisa que deve acontecer para que esse processo seja corretamente é a "ativação" de um dos grupos hidroxila que participa do processo por meio de hidroxila que participa da reação.
Se essa molécula se ligar ao diacilglicerol, é formado o diacilglicerol, o CDP-diacilglicerol é formado (a forma "ativada" do ácido fosfatídico), mas isso também pode acontecer no grupo hidroxila do grupo "cabeça".
No caso da fosfatidilserina, por exemplo, o diabilglicerol é ativado pela condensação da molécula de ácido fosfatídico com uma molécula de trif-citidina citidina (CTP), formando CDP-diacilglicerol e eliminando um pirofosfato.
Se uma molécula de MMP (monofosfato de citidina) se move através de um ataque nucleofílico da serina ou hidroxil hidroxil no carbono 1 do glicerol 3-fosfato. O fosfato produz fosfatidilglicerol.
Ambas as moléculas produzidas dessa maneira servem como precursores de outros lipídios da membrana, que geralmente compartilham rotas biossintéticas uma da outra uma da outra.
- Síntese de colesterol
O colesterol é uma molécula essencial para animais que podem ser sintetizados por suas células, por isso não é essencial na dieta diária. Esta molécula de 27 átomos de carbono é produzida a partir de um precursor: acetato.
Esta molécula complexa é formada a partir de acetil-CoA em quatro estágios principais:
- A condensação de três unidades de acetato para formar o mevalonato, uma molécula intermediária de 6 carbonos (primeiro uma molécula de acetoacetil-coa é formada com duas acetil-coa (enzima tiolase) e depois outro β-hidroxi-β-milglutaril-coA (hmg. COA) (enzima HMG-CoA Syntheta). O Mevalonato é formado a partir do HMG-CoA e graças à enzima hmg-coa redutase.
- MEVALONATO Conversão em unidades de isopreno. Primeiros 3 grupos de fosfato são transferidos de 3 moléculas ATP para Mevalonato. Um dos fosfatos é perdido junto com o grupo carbonil adjacente e é formado
- Polimerização ou condensação de 6 unidades de isopreno de 5 átomos de carbono para formar os átomos de 30 carbonas (uma molécula linear) barbear (uma molécula linear).
- Cicration of the escualia para formar os 4 anéis do núcleo esteróide do colesterol e subsequentes mudanças químicas: oxidações, migração e eliminação de grupos metil, etc., O que produz colesterol.
Referências
- Garrett, r. H., & Grisham, C. M. (2001). Princípios de bioquímica: com um foco humano. Brooks/Cole Publishing Company.
- Murray, r. K., Granner, d. K., Mayes, p. PARA., & Rodwell, V. C. (2014). Bioquímica ilustrada de Harper. McGraw-Hill.
- Nelson, d. eu., Lehninger, a. eu., & Cox, M. M. (2008). Lehninger Principles of Biochemistry. Macmillan.
- Jacquemyn, J., Cascaho, a., & Goodchild, r. E. (2017). Os meandros do retículo endoplasmático - biossíntese lipídica controlada. EMBO Reports, 18 (11), 1905-1921.
- Ohlrogge, J., & Browse, J. (novecentos e noventa e cinco). Biossíntese lipídica. A célula da planta, 7 (7), 957.
- « José López Portillo Biografia, Governo e Contribuições
- Selva tropical nas características do México, localização, alívio, flora, fauna »