

Componente da cadeia transportadora de elétrons, sequência, inibidores
- 3422
- 864
- Ralph Kohler
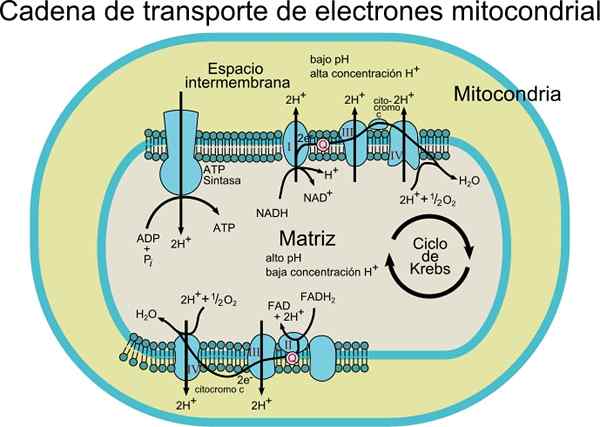
O Cadeia transportadora de elétrons Consiste em um conjunto de moléculas de proteína e coenzima dentro de uma membrana. Como o nome indica, é responsável pelo transporte de elétrons das coenzimas NADH ou FADH2 para o receptor final que é O2 (oxigênio molecular).
Neste processo de transporte, a energia liberada transferindo elétrons de coenzimas para oxigênio molecular por centros redox Unidos para proteínas está associado à produção de energia (ATP). Essa energia é obtida graças ao gradiente de prótons que é gerado na membrana interna mitocondrial.
Este sistema de transporte consiste em vários componentes que podem ser encontrados em pelo menos dois estados de oxidação. Cada um deles é efetivamente reduzido e reoxyd durante o movimento de elétrons de NADH ou FADH2 para O2.
As coenzimas NAD+ e FAD são reduzidas em rotas de oxidação de ácidos graxos e o ciclo do ácido cítrico como resultado da oxidação de vários substratos. Posteriormente, essas coenzimas são oxidadas na cadeia de transporte eletrônico.
Para que o sistema de transporte eletrônico consiste em uma sequência de reações de redução de óxido que estão conectadas entre si.
[TOC]
Componentes de cadeia
Dependendo do tipo de organismo, 3 a 6 componentes podem ser observados constituindo a cadeia de transportadores de elétrons. O processo de transporte de elétrons e a síntese de ATP por fosforilação oxidativa são processos que ocorrem em uma membrana.
No caso de células procarióticas (bactérias aeróbicas), esses processos acontecem associados à membrana plasmática. Nas células eukaryot, ocorre na membrana mitocondrial, de modo que os componentes de transporte de elétrons são encontrados na parte interna da membrana.
https: // giphy.com/gifs/htksbbedquk9z7co5e
Os elétrons são transferidos gradualmente através de quatro complexos que compõem a cadeia de transporte eletrônico.
Cada complexo possui vários componentes proteicos associados a grupos protéticos (componentes não -aminoácidos das proteínas conjugadas) redox, que permitem que seus potenciais de redução aumentem.
Além disso, esse sistema de transporte é composto de várias espécies moleculares, como flavoproteína; coenzima que também chamou Ubiquinona (coq ou uq); vários citocromos como o citocromo B, C, C1, A e A3; Proteínas com Fe-S e grupos de proteínas ligados ao CU. Essas moléculas estão ligadas à membrana, com exceção do citocromo c.
Complexo i
https: // giphy.com/gifs/rlf1rl9gdc6356sj6w
O complexo I chamado NADH coenzima quinona oxidortaduase, ou NADH desidrogenase, consiste em cerca de 45 cadeias polipeptídicas e contém uma molécula de flavina mononucleotídeo (FMN) e oito a nove grupos Fe Fe. Como o nome indica, esse complexo transfere um par de elétrons da nadh coenzyme para coq.
A função do complexo NADH desidrogenase começa com a união de Nadh para o referido complexo ao lado da matriz interna da membrana mitocondrial. Os elétrons são então transportados de NADH para o FMN. Posteriormente, os elétrons passam da flavina reduzida (FMNH2) para as proteínas Fe-S.
O FMNH2 funciona como uma espécie de ponte entre as proteínas NADH e Fe-S, já que esta pode apenas transferir um único elétron, enquanto a coenzima NADH transfere dois, de modo que as flavinas fazem essa transferência de um único elétron graças ao seu redox do semi -cinono.
Pode atendê -lo: monocitopoiese: estágios, características, regulamentaçãoFinalmente, os elétrons são transferidos dos agrupamentos Fe-S para a coenzima Q, que é um transportador de elétrons móveis com uma cauda de isopreneid que o torna hidrofóbico, permitindo atravessar o centro da membrana mitocondrial.
Complexo II
![]()
O complexo II, mais conhecido como succinato desidrogenase, é uma proteína integral da membrana interna mitocondrial e é uma enzima que intervém no ciclo do ácido cítrico.
Este complexo consiste em duas subunidades hidrofílicas e duas hidrofóbicas com grupos Hemo B que fornecem o local de ligação do COQ, além de uma flavoproteína e uma proteína com Fe-S.
No ciclo do ácido cítrico (Krebs ou ciclo de ácido tricarboxílico), o succinato é convertido em fumarato por succinato desidrogenase, reduzindo a coenzima FAD para FADH2. A partir desta última coenzima, os elétrons são transferidos para os centros Fe-S que, por sua vez, os transferem para o COQ.
Durante as reações desta transferência de elétrons, o potencial redox padrão é muito baixo, o que impede que a energia livre necessária seja liberada para sintetizar o ATP.
Isso significa que o complexo II é o único complexo na cadeia de transporte de elétrons, incapaz de fornecer energia para a síntese de ATP. No entanto, esse complexo é fundamental no processo, pois transfere elétrons do FADH2 para o restante da cadeia.
Complexo iii
https: // giphy.com/gifs/cjg0raiuizjds2hkyu
O complexo III, o complexo do citocromo BC1 ou o citocromo c redutase de coq, transfere os elétrons da coenzima reduzida para o citocromo c. Esta transferência ocorre através de uma única estrada redox, que é conhecida como Ciclo Q.
Este complexo consiste em uma proteína com Fe-S e três citocromos diferentes, nos quais o átomo de ferro localizado no grupo Hemo varia ciclicamente entre os estados reduzidos (Fe2+) e oxidado (Fe3+).
Os citocromos são hemoproteínas de transporte eletrônico, que têm atividade redox. Eles estão presentes em todos os organismos, com exceção de alguns anaerobes obrigatórios.
Essas proteínas têm grupos Hemo que alternam entre dois estados de oxidação (Fe2+ e Fe3+). O citocromo c é um transportador de elétrons móveis fracamente associado à membrana interna das mitocôndrias.
Os citocromos encontrados neste complexo são os citocromos B, C e A, os 3 são proteínas redox ativas com grupos, temos características diferentes, que alternam seus estados de oxidação entre Fe2+ e Fe3+.
O citocromo c é uma proteína da membrana periférica que funciona como um "transporte" de elétron com citocromo c1 e com o complexo IV.
IV complexo
O citocromo c e o2 são os receptores finais de elétrons derivados da oxidação do material orgânico; portanto, o complexo IV ou citocromo c oxidase é a enzima terminal no processo de transporte eletrônico. Isso aceita elétrons do citocromo c e os transfere para a redução de O2.
A função complexa é catalisar oxidações de um elétron das quatro moléculas consecutivas do citocromo C reduzido, ou seja, reduz simultaneamente quatro elétrons de uma molécula de O2, finalmente produzindo duas moléculas de H2O.
Pode atendê -lo: tecido de revestimento epitelialSequência de transporte de elétrons
Os elétrons são transferidos dos complexos I e II para o complexo III, graças à coenzima q, e a partir daí passam para o complexo IV através do citocromo c. Quando os elétrons passam por esses quatro complexos, o potencial de redução aumenta, liberando energia, que é então usada para síntese de ATP.
No total, a transferência de alguns elétrons causa a translocação de 10 prótons através da membrana; quatro nos complexos I e IV e dois dos complexos III.
Nadh Descidrogenasa
Esta enzima catalisa a oxidação da coenzima Nadh pela coenzima que. Os elétrons se movem do NADH para o FMN que está ligado à cauda hidrofílica do complexo i. Fe-S Groupings Transfer para elétrons em um momento. Esses grupos Fe-S reduzem ao coq, que é incorporado na membrana, ao ubiquinol (CoQ reduzido).
Durante a transferência de elétrons para o COQ, quatro prótons são transferidos pela membrana interna, para o espaço intermembranal. O mecanismo pelo qual esses prótons são translocados envolve proteínas localizadas na cauda hidrofóbica do complexo i.
O processo de transferência de elétrons nesta etapa libera energia livre, especificamente -16,6 kcal/mol.
Coq-citocromo c redutase e o ciclo Q
A coenzima que é oxidada pelo citocromo c, em uma reação catalisada por esta coenzima. A oxidação do ubiquinol (CoQ reduzida) ocorre em um determinado local do complexo (qo ou local de oxidação) na membrana mitocondrial, transferindo dois elétrons, um para a proteína com os grupos Fe-S e outro para os grupos Hemo.
No ciclo Q, a oxidação de Coq produz semi -quinona, que é onde os elétrons são transferidos para os grupos Hemo B1 e BH. Enquanto essa transferência de elétrons ocorre, um segundo coq oxida no local de Qo, repetindo o ciclo.
Este ciclo causa a transferência de dois elétrons e, por sua vez.
Citocroma C oxidase
Esta enzima (complexo IV) catalisa a oxidação do citocromo c (reduzido) por O2, que é o aceitador final dos elétrons. Esta transferência produz uma molécula H2O para cada par de elétrons transferidos, além da translocação de prótons através da membrana.
Os elétrons se movem de um para um, do citocromo C reduzido para um par de íons CUA, depois passam para um grupo hemo e finalmente alcançam o centro binuclear do complexo que contém íons Cub e Hemo A3, onde a transferência de quatro elétrons ocorre para oxigênio.
No complexo IV, os elementos transferem os elétrons de um por um, de modo que o O2 é reduzido gradualmente, de modo que a liberação de alguns compostos tóxicos, como superóxido, peróxido de hidrogênio ou radicais hidroxila, não ocorre.
A energia liberada nesta fase corresponde a -32 kcal/mol. O gradiente eletroquímico gerado durante o processo de transferência e as mudanças de energia (ΔE) causadas por alguns elétrons ao passar pelos quatro complexos corresponde, em cada estágio, à energia livre necessária para a produção de uma molécula ATP.
Pode atendê -lo: teoria dobrável da membranaGrande desidrogenase
Como mencionado, esse complexo tem a única, mas importante, função de introduzir os elétrons FADH2 do ciclo do ácido cítrico para a cadeia de transporte de elétrons.
Esta enzima catalisa a oxidação da coenzima FADH2 pela coenzima Q (oxidada). No ciclo do ácido cítrico quando o succinato ao fumarato, dois elétrons e dois prótons são transferidos para a moda. Posteriormente, o FADH2 transfere esses elétrons até coq através dos centros Fe-S do complexo.
Finalmente, a partir do coq, os elétrons são transferidos para o complexo III, seguindo as etapas descritas acima.
Os complexos de cadeia são independentes
Os quatro complexos que compõem a cadeia de transporte eletrônico são independentes, ou seja, eles são de forma independente e operam na membrana interna mitochcondrial, e o movimento de cada um deles na membrana não depende ou está ligado aos outros complexos.
Os complexos I e II se movem na membrana transferindo seus elétrons para o COQ que também se espalha na membrana e os transfere para o complexo III, de onde os elétrons passam para o citocromo c que também são móveis na membrana e deposita elétrons no complexo IV.
Inibidores da cadeia de transporte eletrônico
Na cadeia de transporte eletrônico, alguns inibidores específicos agem em seu processo. A rotenona é um inseticida comumente usado que é estequiometricamente unido ao complexo I, evitando a redução do COQ.
Alguns barbitúricos tipo medicamentos como Piericidina e Amytal, inibem o complexo I, interferindo na transferência de elétrons dos grupos Fe-S para o COQ.
No complexo II, alguns compostos como teoiltrifluoroacetona e malonato atuam como inibidores competitivos com succinato, evitando sua oxidação e, por sua vez, a transferência de elétrons para a moda.
Alguns antibióticos como mixotiazol e estigmatellina se ligam ao local Q do CoQ, inibindo a transferência de elétrons da coenzima Q para os centros Fe-S das proteínas.
Cianeto, azida (N3-), ácido sulfúrico e monóxido de carbono inibem o complexo IV. Esses compostos se ligam aos grupos Hemo, evitando a transferência de elétrons para o centro binuclear complexo ou de oxigênio (O2) (O2).
Quando a cadeia transportadora de elétrons é inibida, a produção de energia é interrompida devido à fosforilação oxidativa, causando danos graves e até a morte do organismo.
Referências
- Alberts, b., Bray, d., Hopkin, k., Johnson, a., Lewis, J., Raff, m., Roberts, k. & Walter, P. (2004). Biologia celular essencial. Nova York: Garland Science. 2ª edição.
- Cooper, g. M., Hausman, r. E. & Wright, n. (2010). A célula. (pp. 397-402). Ed. Marbán.
- Devlin, t. M. (1992). Livro de Bioquímica: com correlações clínicas. John Wiley & Sons, Inc.
- Garrett, r. H., & Grisham, C. M. (2008). Bioquímica. Ed. Thomson Brooks/Cole.
- Rawn, j. D. (1989). Bioquímica (no. 577.1 cru). Ed. Interamericano-McGraw-Hill
- Voet, d., & Voet, J. G. (2006). Bioquímica. Ed. Pan -American Medical.
- « Características de Anguila Elétrica, Habitat, Reprodução
- Características de Biozoos, Morfologia, Reprodução, Nutrição »